


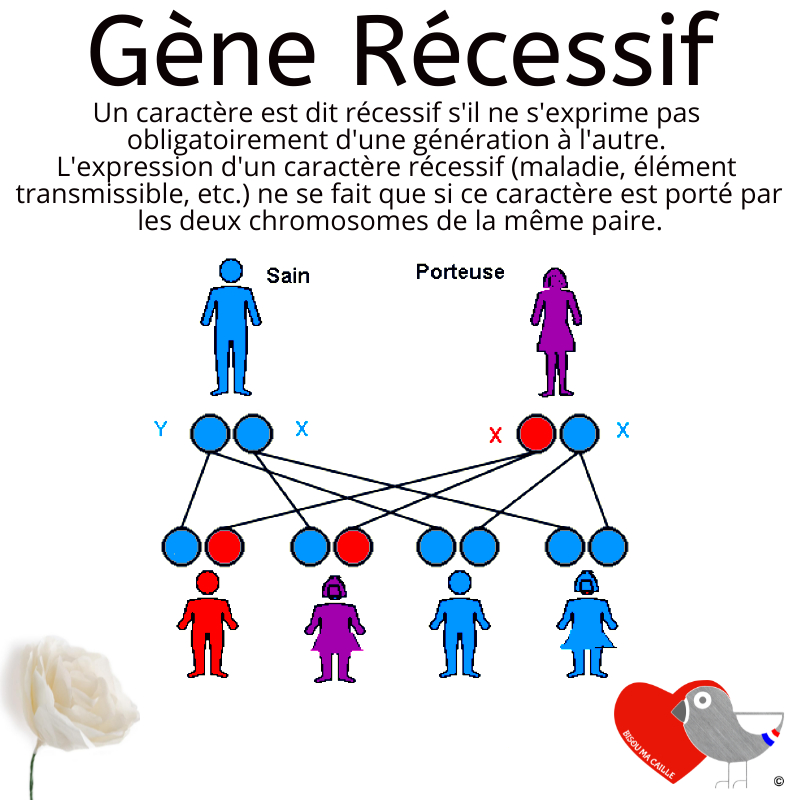
Les plumages auto-sexués ou liés au sexe sont utiles pour différencier les mâles et les femelles à l’éclosion, en particulier pour ceux qui préfèrent n’élever que des oiseaux femelles à des fins de ponte. Les plumages à sexage automatique sont des plumages qui se présentent différemment chez les mâles et les femelles à l’éclosion, et la progéniture continuera à présenter ces différences de plumage mâle/femelle génération après génération. Les descendants auto-sexués sont de race pure.
En revanche, les plumages liés au sexe se présenteront différemment chez les mâles et les femelles à l’éclosion pendant une génération en fonction de la génétique du plumage des parents, mais la progéniture ne continuera pas à présenter la même différenciation de plumage mâle / femelle lorsqu’elle est élevée.
La reproduction liée au sexe chez les cailles implique généralement des mutations sur le chromosome Z. Contrairement aux humains, où les mâles ont un chromosome X et un Y et les femelles ont deux chromosomes X, chez la caille, les mâles ont deux chromosomes Z et les femelles ont un chromosome Z et un W. Le chromosome W chez les cailles femelles est extrêmement court et est donc moins susceptible de porter des mutations, similaires au chromosome Y chez les mâles humains.
La reproduction de plumages liés au sexe chez la caille japonaise est le plus souvent basée sur la mutation Roux (BR*R) qui est située sur le chromosome sexuel (Z). La reproduction liée au sexe est accomplie lorsque des mâles homozygotes pour la mutation Roux sont accouplés à des femelles sans la mutation, ce qui donne une génération de descendants qui peuvent être identifiés à l’éclosion comme des femelles Roux hémizygotes et des mâles porteurs qui se présentent comme un phénotype non roux.
En clair, un Locus de Gène contient 2 Allèles.
Hétérozygote = L’Allèle n’est présent qu’une seule fois
Homozygote = L’Allèle est présent deux fois
AD = Autosomique dominant sur l’allèle de type Sauvage
AR = Autosomique récessif à l’allèle de type Sauvage
SR = Lié au sexe récessif à l’allèle de type Sauvage.
Presque tout les AD sont essentiellement incomplètement dominants.

Continuation de notre guide sur les Mutations dans la Caille du Japon
Les Mutations de couleurs
Une des dernières mutation de couleur dont l’ADN as été séquencé provient de recherches menées ici même en France!
La couleur « Rouillé » “Rusty” as été séquencé en 2005 par Francis Minvielle, Département Génétique et Diversité Animales, Institut National de la Recherche Agronomique, au Centre de Jouy, France.
Générations Filiales: générations nommées dans un croisement génomique, l’appariement d’origine étant la génération F0, leur progéniture étant la génération F1, et chaque génération suivante étant numérotée F2, F3, F4 etc.
Pure race: descendance homozygote qui est le résultat de parents reproducteurs homozygotes pour un allèle spécifique.
Hybride: descendance hétérozygote issue de la reproduction de parents dissemblables.
Gène: unité héréditaire constituée de paires de bases d’ADN à un emplacement spécifique sur un chromosome.
Phénotype: Les caractéristiques observables.
Génotype: La construction génétique (non observable) d’un individu.
Allèle: Forme de mutation. Une ou plusieurs séquences d’ADN se produisant à un locus de gène particulier.
Locus: Localisation d’un allèle.
Mode d’hérédité: La manière dont un trait génétique ou une mutation est hérité.
Mutation: Une version modifiée du matériel génétique résultant en un nouvel allèle.
Héritage: La manière dont le matériel génétique est transmis des parents à la progéniture.
Allèles Dominants: Hérédité autosomique dominante, la mutation s’exprime lorsqu’un variant est présent dans un seul allèle (copie) d’un gène donné. Masquera ou s’exprimera sur d’autres couleurs.
Allèles récessifs: Une variation d’un gène qui n’affecte pas un phénotype lorsqu’un allèle dominant est présent. Hérédité autosomique récessive, la mutation s’exprime lorsqu’un variant est présent sur les deux allèles (copies) d’un gène donné. Mutation exprimée lorsqu’une copie est héritée de chaque parent.
Allèles co-dominants: Une variation d’un gène qui produira un effet dominant dans une partie d’un organisme, tandis qu’un autre allèle produit un effet différent dans une autre partie de l’organisme.
Autosomique Incomplète dominante: 1 copie change de couleur, 2 copies change à nouveau de couleur.
Dominant autosomique incomplet: L’hérédité dominante incomplète se produit lorsqu’une copie n’exprime que partiellement et ne masquera pas complètement les autres (hétérozygote).Deux copies la modifient à nouveau (homozygote).
Autosomique incomplet récessif: 1 copie peut changer de couleur, mais pas toujours, 2 copies, 1 de chaque parent, la modifie à nouveau.
Hétérozygote: Présence de deux allèles différents à un locus de gène particulier.
Homozygote: Présence de deux allèles identiques à un locus de gène particulier.
Chromosomes: Structure filiforme contenant du matériel génétique. (Les cailles ont 39 paires de chromosomes.)
Autosome: Tout chromosome autre qu’un chromosome sexuel. (Les cailles ont 38 paires d’autosomes.)
Chromosomes sexuels: L’un des deux types de chromosomes impliqués dans la détermination du sexe d’un organisme. Chez la caille, les chromosomes sexuels sont désignés par Z et W. Les mâles sont homomorphes (ZZ) et les femelles sont hétéromorphes (ZW).
Épistasie: scénario dans lequel la fonction d’un allèle dépend d’un ou plusieurs allèles hérités indépendamment.
Hypostase: scénario dans lequel la fonction d’un allèle est inhibée par un ou des allèles épistatiques.
Cryptomères: Un allèle dominant qui est masqué par la fonction d’un ou plusieurs allèles hérités indépendamment.

Introduction


BASE PRINCIPALE 1 : Sauvage (Commun) (Locus: MC1R)
Nom Anglais ISO: “Pharaoh”

C’est le motif de plumage sur lequel toutes les autres couleurs sont construites.
L’appellation en Anglais est: “Pharaoh”, “Wild”, ou “Pharaoh Wild”.
Une lignée pure et avec une haute qualité de plumage de ce coloris (c’est-à-dire que la lignée se reproduit fidèlement de génération en génération sans aucune variation au fil des génération) est essentielle pour tout programme d’élevage de couleurs, et devient votre base “maison” pour tout croisement qui requiert cette base.
Les oiseaux de type Sauvage/Commun sont presque identiques aux Cailles Japonaises Sauvages, bien qu’ils puissent varier considérablement en taille et en tempérament.
Lorsque nous discutons d’autres mutations du plumage, elles seront généralement comparées à ce type Sauvage, et leur héritage sera défini par rapport au plumage de type Sauvage, qui se compose de diverses nuances de brun et de blanc, et est sexuellement dismorphique.
Les mâles affichent des facettes et un plumage ventral radicalement différent des femelles.
Vers l’âge de trois semaines, les mâles développent un plumage ventral de couleur cuivre distinct, tandis que les femelles conservent un plumage ventral de couleur crème avec une plume centrale vaine qui apparaît comme des taches noires déconnectées sur la majorité du plumage ventral.
Lorsqu’il n’y a pas de gènes récessifs/cachés présents, Sauvage x Sauvage produira une progéniture 100 % Sauvage.
Pour les croisements:
Sauvage x Sauvage = 100% Sauvage



BASE PRINCIPALE 2 : Brun Étendu (Locus : MC1R)
Nom Anglais ISO: “Extended Brown” Mutation
Tibétain:
Le Tibétain se présente lorsqu’un oiseau est homozygote pour la mutation “Extended Brown”. Le Tibétain est un oiseau brun foncé, sans plumes striées et avec un plumage foncé sur tout le corps. La peau des pattes d’un oiseau tibétain sera brun foncé et s’étendra souvent à certaines parties des pattes. Généralement, les orteils resteront roses, bien qu’une quantité beaucoup plus petite de la surface restera rose par rapport à une Rosetta. Prolonge la pigmentation noire et marron foncé sur tout le corps. Il n’y a pas de différence entre le plumage d’un mâle et celui d’une femelle.
Rosetta:
Le Rosetta se présente lorsqu’un oiseau est hétérozygote pour la mutation “Extended Brown”. Rosetta est un oiseau brun foncé, avec des stries claires et un plumage foncé sur tout le corps. La peau des pattes d’un oiseau Rosetta sera brun clair et s’étendra souvent à certaines parties des pieds. Généralement, les orteils resteront roses. Prolonge la pigmentation noire et brun foncé sur tout le corps avec des barres dorées au centre des plumes. Il n’y a pas de différence entre le plumage d’un mâle et celui d’une femelle.
Notes: Le Rosetta est très similaire en apparence avec la couleur du Tibétain, mais se différencie sur les épingles en or qui descendent au centre des plumes. Cela aide à distinguer les Rosetta des Tibétains, qui eux sont homozygotes Marron Étendu (EB).
N’est pas sexable par le motif du plumage.
Pour les croisements:
Sauvage x Tibétain = 100% Rosetta
Sauvage x Rosetta = 50% Sauvage, 50% Rosetta



BASE PRINCIPALE 3 : Fauve (Isabelle) (Locus : ASIP)
Nom Anglais ISO: “Fawn”
Fauve / Italien / Isabelle:
Le Fauve / Italien / Isabelle se présente lorsqu’un oiseau est hétérozygote pour la mutation “Fauve” (“Fawn en Anglais).
Le motif du plumage et la couleur changent, donnant un oiseau jaune avec des quantités approximativement égales de noir et de rouge sur le plumage.
Le motif est assez cohérent, avec des barres noires diagonales en forme de chevron de chaque côté des plumes dorsales, précédées de taches rouges, et un centre et une base jaunes.
À l’âge adulte, les mâles Fauve ont un plumage jaune-beige crémeux, de plus grandes marques sombres sur le dos et un visage brun. Les femelles adultes seront de couleur jaune-beige crémeux avec de plus grandes marques noires sur la tête et le dos. Les femelles Fauve auront également des taches sombres sur la poitrine.
Manchurian:
Manchurian ou “Golden Manchurian” se présente lorsqu’un oiseau est homozygote pour la mutation “Fawn” (“Fawn en Anglais).
Le motif et la couleur du plumage changeront tous les deux, donnant un oiseau jaune avec de petites taches noires non connectées sur le plumage ventral.
À l’âge adulte, les mâles homozygotes Manchurian ont un plumage jaune-beige crémeux, très peu de marques sur le dos et un masque facial et un collier de couleur brun rougeâtre et blanc. Les femelles adultes seront de couleur crème jaune-beige avec des marques noires sur la tête et le dos.
Pour les croisements:
Sauvage x Fauve = 50% Sauvage, 50% Fauve
Sauvage x Manchurian = 100% Fauve
BASE PRINCIPALE 4 : Pansy (Locus: MC1R)
Nom Anglais ISO: “Pansy”
Le “Pansy”, également connue sous le nom de “Redhead” ou “Rotkopf”, est un phénotype causé par une mutation récessive sur le locus brun étendu. Il fait éclaircir le plumage à la base de l’oiseau en un brun jaunâtre et modifie la striation des plumes en barrant. Chaque plume a une barre noire épaisse, une barre marron clair et une barre blanche étroite sur le bord extérieur de la plume. Cela donne à l’oiseau un aspect marbré sur tout le corps. Les mâles ont un masque rouge qui les distingue facilement des poules.
Corps principalement blanc avec des marques irrégulières de couleur noire et rouille. Les mâles ont une tête de couleur rouge rouille.
Pour les croisements:
Sauvage x Pansy = 100% Sauvage (avec un % de porteurs de gènes Pansy)


BASE PRINCIPALE 5 : Sparkly (Locus: MC1R)
Nom Anglais ISO: “Sparkly”
Le “Sparkly” se présente lorsqu’un oiseau est homozygote pour la mutation Sparkly. Les stries du plumage dorsal sont remplacées par des barres sombres et de fines barres claires distinctes dont une sur le bord extérieur des plumes, tandis que le plumage ventral devient panaché et sombre, même chez les mâles. Les parties sombres des plumes sont exagérées et le motif est cohérent.
Grosses marques noires et brunes. Parfois identique aux poussins Sauvage selon la variété de Sparkly qui est croisée.
Heterozygous Sparkly crée une poitrine pleine de taches sur les Femelles (jusqu’aux pattes) et une poitrine mouchetée / à motifs sur les Males également.
Pour les croisements:

BASE PRINCIPALE 6 : Calico (Locus: INCONNU)
Nom Anglais ISO: “Calico”
Le “Calico” est une mutation qui provoque un éclaircissement du plumage et une modification du motif du plumage. La mutation éclaircit le plumage ventral et facial, ce qui rend légèrement plus difficile la distinction des mâles des femelles. Plus particulièrement, il exagère les stries sur le plumage ventral, donnant aux oiseaux Calico un motif elliptique unique sur le dos.
Pour les croisements:
Sauvage x Calico Hétérozygote = 50% Sauvage, 50% Calico Hétérozygote
Sauvage x Calico Homozygote = 100% Calico Hétérozygote
BASE PRINCIPALE 7 : Noir Récessif (Locus : MC1R)
Nom Anglais ISO: “Recessive Black”
La mutation récessive du plumage noir chez la Caille Japonaise est contrôlée par un gène autosomique récessif (rb) et présente un phénotype brun noirâtre à l’état homozygote récessif (rb/rb). Un test d’allélisme a montré que le type sauvage et les mutations pour le jaune, le fauve-2 et le noir récessif chez la Caille Japonaise étaient des allèles multiples (*N, *Y, *F2 et *RB) au même locus Y et que la relation de dominance était YF2 > YY > YN > YRB.
Pour les croisements:

Ces Gènes de Dilution (“Diluter Genes” en Anglais) modifient la palette de couleurs du plumage, mais pas le motif de base sous-jacent de l’oiseau.
L’un des aspects fascinants de la Caille du Japon est la diversité de ses couleurs de plumage, qui sont principalement déterminées par différents gènes. Parmi ceux-ci, les gènes de dilution jouent un rôle clé dans la modification de l’intensité des couleurs.
Les gènes de dilution sont des gènes qui modifient l’apparence des autres gènes responsables de la couleur du plumage. En termes simples, ils agissent comme des régulateurs de l’intensité de la couleur, permettant une variation dans la palette de couleurs observée chez la Caille du Japon. Ces gènes n’influencent pas directement la couleur du plumage, mais modifient la manière dont les autres gènes s’expriment, créant ainsi des nuances plus claires ou plus foncées.
Il existe plusieurs gènes de dilution connus chez la Caille du Japon. Parmi eux, on peut citer le gène “dilute” (d), le gène “pale” (p) et le gène “silver” (s). Chacun de ces gènes a un effet spécifique sur le plumage de la Caille.
-
- Le gène “dilute” (d) est responsable de la réduction de l’intensité des pigments, ce qui donne un aspect dilué aux couleurs. Les Cailles portant ce gène auront des plumes plus claires que celles sans le gène.
-
- Le gène “pale” (p) provoque une diminution de la pigmentation des plumes, ce qui entraîne un plumage plus pâle. Les Cailles avec ce gène présenteront des plumes d’une couleur crème ou pâle, plutôt que d’une couleur plus foncée.
-
- Enfin, le gène “silver” (s) affecte la distribution de l’eumélanine, un pigment responsable de la couleur sombre du plumage. Les Cailles portant le gène “silver” auront des plumes d’un gris argenté, ainsi qu’une réduction de la pigmentation noire.
Les gènes de dilution chez la caille japonaise sont essentiels pour comprendre et apprécier la diversité des couleurs de plumage présente chez cette espèce. L’étude de ces gènes offre des perspectives intéressantes sur la génétique, l’élevage et la sélection de ces oiseaux pour différents usages et appréciations esthétiques.



“Argenté” (Silver en Anglais)
“Argenté” est homozygote et a un facteur létal, ce qui peut éventuellement causer des problèmes de mortalité élevées si trop de “in-breeding” as été fait sur cette souche. Il a été réduit par la reproduction avec une génétique de dilution plus variée, mais il est probable qu’il se reproduise si vous reproduisez trop longtemps ensemble de l’Argenté avec de l’Argenté.
Crédit Photo: Cora Guindon, USA. Merci ! 🙂
“Andalousie” (Andalusian en Anglais)
Un oiseau Andalousie hétérozygote peut avoir des primaires blanches et un motif de Pied d’Aile Blanche ou être principalement Argenté avec des tons bruns. Il peut avoir des taches de plumage non dilué ou des plumes solitaires réparties au hasard sur le corps. L’Andalousie homozygote apparaîtra blanc cassé avec des yeux foncés. Semblable à l’Argenté, la mutation Andalousie interagit avec presque toutes les autres combinaisons de mutations, produisant un nombre infini de phénotypes.
“Bleu” (Blue/Blau en Anglais)
Voir section dédiée au Bleu ci-dessous.
“Fee”
Mutation qui change le plumage brun en blanc, peut être homozygote ou hétérozygote.
“Fee” est une mutation de dilution de couleur qui laisse le pigment noir et gris dans le plumage mais dilue le pigment brun.
Pour les différents types de Fee, vous avez besoin d’au moins un oiseau parent spécifique dans une variation de Fee, pour intégrer la mutation de Fee dans la lignée sanguine.
Le gène Fee est un allèle dominant: si vous avez un Male Fee homozygote, vous pouvez le croiser avec plusieurs Femelles Sauvages et vous devriez avoir dans la génération F1 100% Fee, qui seront hétérozygotes.
Après la génération F1, vous obtiendrez (statistiquement) dans les générations suivantes F2, F3, F… : 25% de non-Fee, 50% de Fee hétérozygotes et 25% de Fee homozygotes.
Comme le gène des Fee est une paire d’allèles distincte du gène du motif de couleur principal, le motif sous la dilution des Fee est toujours dans son propre héritage dominant, récessif ou intermédiaire…
Transforme en Blanc les couleurs de base de type Sauvage et Fauve. Si sous forme homozygote, vous obtenez le double de la dilution Blanche !

“Roux”
Mutation qui change le plumage noir en un brun châtain chaud, récessif lié au sexe.
“Sandy”
Pour les croisements:

Exemples de Dilutions:
“Sauvage” dilué avec du “Fee”:
Falb Fee
“Falb Fee” est le nom commun du croisement de Sauvage avec Fee, que l’oiseau soit hétérozygote ou hétérozygote pour Fee.
L’oiseau se présente comme une version en niveaux de gris de l’oiseau non dilué. Sous forme hétérozygote, l’oiseau est partiellement dilué, et sous forme homozygote, il sera presque entièrement dilué. Seuls les mâles montreront un soupçon de plumage facial non dilué, tandis que le reste de l’oiseau est noir, blanc et gris. Une Caille Femelle idéale aura une absence totale de plumage brun.
“Fauve” dilué avec du “Fee”:
Pearl Fee
“Pearl Fee” ou “Italian Fee” est le nom commun du croisement de Fauve avec Fee, que l’oiseau soit hétérozygote ou hétérozygote pour Fee et/ou Fauve.
Il est difficile d’identifier les poussins Pearl Fee des poussins Fauve standard. Cela n’est que légèrement plus facile si vous avez les deux couleurs dans le même lot. Les différences sont extrêmement subtiles, mais les poussins Fauve ont tendance à être plus dorés tandis que les poussins perlés sont d’un jaune plus clair partout. Les poussins perlés semblent également avoir de fines rayures «pointillées» sur le dos ou des taches supplémentaires, tandis que les poussins Fauve ont des rayures assez uniformes. Les poussins perlés peuvent aussi avoir des points sur le visage près de leurs oreilles, contrairement au poussin Fauve qui peut avoir un eye-liner classique.
“Brun Étendu” dilué avec du “Fee”:
Grau Fee
“Grau Fee” est le nom commun de Rosetta (Brun Étendu hétérozygote) ou Tibétain (Brun Étendu homozygote), que l’oiseau soit hétérozygote ou homozygote pour Fee.
“Pansy” dilué avec du “Fee”:
Pansy Fee
“Pansy Fee” est le nom commun du Pansy homozygote, que l’oiseau soit hétérozygote ou homozygote pour Fee.
Pearl Fee Sparkly (si dilué par du Fauve)
“Pearl Fee Sparkly” est le nom commun de Sparkly hétérozygote sur Fauve avec Fee, que l’oiseau soit hétérozygote ou homozygote pour Fee.
Falb Fee Sparkly (si dilué par du Sauvage)
“Falb Fee Sparkly” est le nom commun de Sparkly hétérozygote sur Sauvage avec Fee, que l’oiseau soit hétérozygote ou homozygote pour Fee.
“Sauvage” dilué avec du “Roux”:
Égyptien
“Égyptien” (“Egyptian” en Anglais) est le nom commun du croisement du Sauvage avec un porteur/se Roux qui est visuellement présent sur l’animal. Hétérozygote chez les Femelles et homozygote chez les Males. Si un Male Sauvage n’a qu’un seul exemplaire de Roux (qui ne serait pas visible au-delà d’une légère amélioration de la pigmentation), il serait communément appelé “Porteur Roux” ou “Roux Carrier” en Anglais.
Motif de plumes sauvages grisâtres et brun clair couvrant tout le corps. Les mâles ont une poitrine solide et crémeuse, tandis que les femelles ont des taches brun clair sur la poitrine.
“Fauve” dilué avec du “Roux”:
Ambre d’Automne
Ambre d’Automne (“Autumn Amber” en Anglais) est le nom commun du croisement du Fauve (Fauve hétérozygote) avec un porteur/se Roux qui est visuellement présent sur l’animal. Hétérozygote chez les Femelles et homozygote chez les Males. Si un Male Fauve n’a qu’un seul exemplaire de Roux (qui ne serait pas visible au-delà d’une légère amélioration de la pigmentation), il serait communément appelé “Porteur Roux” ou “Roux Carrier” en Anglais. L’Ambre d’Automne portent les marques des motifs de plumes dorées de la Mandchourie et de la Fauve, mais la base beige crémeuse prend davantage une couleur crème orange et les marques noires deviennent brunes.
Ambre de Mandchourie
“Ambre de Mandchourie” (“Amber Mandchurian” en Anglais) est le nom commun du Mandchourien (Fauve homozygote) avec un porteur/se Roux qui est visuellement présent sur l’animal. Hétérozygote chez les Femelles et homozygote chez les Males. Si un Male Fauve n’a qu’un seul exemplaire de Roux (qui ne serait pas visible au-delà d’une légère amélioration de la pigmentation), il serait communément appelé “Porteur Roux” ou “Roux Carrier” en Anglais.
“Rosetta” dilué avec du “Roux”:
Scarlett
“Scarlett” (Nom Anglais) est le nom commun du croisement du Rosetta (Brun Étendu hétérozygote) avec un porteur/se Roux qui est visuellement présent sur l’animal. Hétérozygote chez les Femelles et homozygote chez les Males. Si un mâle Rosetta n’a qu’un seul exemplaire de Roux (qui ne serait pas visible au-delà d’une légère amélioration de la pigmentation), il serait communément appelé “Porteur Roux” ou “Roux Carrier” en Anglais.
“Tibétain” dilué avec du “Roux”:
Range (ou Range Rouge en Anglais)
“Range” (Nom Anglais) est le nom commun du croisement du Tibétain (Brun Étendu homozygote) avec un porteur/se Roux qui est visuellement présent sur l’animal. Hétérozygote chez les Femelles et homozygote chez les Males. Si un Male Tibétain n’a qu’un seul exemplaire de Roux (qui ne serait pas visible au-delà d’une légère amélioration de la pigmentation), il serait communément appelé “Porteur Roux” ou “Roux Carrier” en Anglais.
Pour les croisements:
Sauvage x Sparkly Heterozygote = 50% Sauvage, 50% Sparkly Heterozygote
Sauvage x Sparkly Homozygote = 100% Sparkly Heterozygote
Sauvage x Égyptien = récessif lié au sexe
Sauvage x Range = récessif lié au sexe
Sauvage x Scarlett = récessif lié au sexe
Sauvage x Ambre d’Automne = récessif lié au sexe
Sauvage x Ambre de Manchurian = récessif lié au sexe
Sauvage x Falb Fee Hétérozygote = 50% Sauvage, 50% Falb Fee Heterozygote
Sauvage x Falb Fee Homozygote = 100% Falb Fee Heterozygote
Sauvage x Grau Fee Hétérozygote = 50% Sauvage, 50% Grau Fee Hétérozygote
Sauvage x Grau Fee Homozygote = 100% Grau Fee Hétérozygote

Motifs de plumage supplémentaires
(en Europe, États-Unis et dans d’autres régions du monde)
Tuxedo
Voir section dédiée au Tuxedo ci-dessous.
Pied d’Aile Blanche (“White Wing Pied” en Anglais)
Le “Pied d’Aile Blanche” Homozygote se présente comme un oiseau au corps blanc, avec une tête et un cou blancs, et un plumage coloré sous-jacent sur tout le dos et la queue. Un spécimen idéal n’aura aucune couleur sur la tête et les ailes. Un oiseau portant une copie de White Wing Pied aura des bouts d’ailes et des plumes primaires blanches.
Ailes et ventre d’un blanc pur, avec une barre de couleur/motif de plumes en forme de selle (comme Sauvage) sur le dos et l’arrière de la tête.
Pied Progressif (“Progressive Pied” en Anglais)
Panda
Panda se présente comme un oiseau blanc avec deux grandes taches montrant le plumage sous-jacent. La première tache est à l’arrière de la tête, semblable à un pointillé blanc, mais plus grande. La seconde est sur le bas du dos. Les oiseaux blancs pointillés peuvent aussi parfois avoir un plumage coloré sur le bas du dos, mais pas aussi grand et cohérent que Panda.
Contrairement au Blanc Pointillé, chez le mutant Panda, les plumes des taches sombres sont similaires au type sauvage à la fois en couleur et en marques (Tsudzuki et al, 1993).
Le Panda présente un plumage blanc avec des taches sombres sur la tête, les joues, le dos, les ailes et la queue.

Blanc Anglais (alias “Blanc pointillé”, “Blanc Tacheté”, ou “English White” et/ou “Dotted White” en Anglais)
Parfois appelés “English White”, les bons spécimens sont complètement blancs, sans marquages discernables, sauf sur la tête. Le bec et les pattes sont brun rosâtre et les yeux sont brun foncé.
Il est courant d’avoir des taches noires sur la tête et le dos.
Les éleveurs qui visent des spécimens tout blancs peuvent élever cela avec une reproduction sélective prudente, mais méfiez-vous! Le blanc est considéré comme un gène récessif, mais ces dernières années un gène albinos incomplètement dominant a également été signalé.
Le mâle et la femelle sont identiques, bien que la femelle soit plus grande.
N’est pas sexable par le motif du plumage.
(Tsudzuki et al, 1992) : Il s’agit d’une mutation autosomique récessive de la couleur du plumage découverte chez le poussin mutant ayant des plumules de duvet jaune crème avec une tache brune sur la tête et/ou le dos des poussins mutants n’ayant pas de tache colorée. Les plumules jaune crème et brunes du poussin remplacent respectivement les plumes blanches et foncées à l’âge adulte. Dans les plumes foncées, toute la pigmentation est diluée et la ligne de tige de couleur paille de blé est plus étroite que le type sauvage. De plus, les barres transversales dans les plumes individuelles sont complètement absentes. Le pointillé blanc est un allèle récessif inférieur au locus panda (s) (Mizutani et al, 1974; Tsudzuki et al, 1993). Le panda est également un mutant qui présente un plumage blanc avec des taches sombres sur la tête, les joues, le dos, les ailes et la queue. Contrairement au blanc pointillé, chez le mutant panda, les plumes des taches sombres sont similaires au type sauvage à la fois en couleur et en marques (Tsudzuki et al, 1993).
Le Blanc pur a généralement des marques brun foncé à l’arrière de la tête. Le blanc pur croisé avec du blanc pur donnera 25 à 50 % de blanc pur.
Splash
Découvert en Australie, et pas encore disponible en Europe.
Pour les croisements:
Sauvage x Blanc Anglais = 100% Tuxedo Sauvage
Blanc Anglais x Blanc Anglais = 25-50% Blanc Anglais, 50-75% Sauvage/Autres

Gènes de Dilution supplémentaires
(en Europe, États-Unis et dans d’autres régions du monde)
Les gènes de dilution qui sont les plus courants, “Bleu” et “Argent”, modifient la palette de couleurs du plumage, mais pas le motif de base sous-jacent de l’oiseau.

Bleu
Bleu (anciennement Lavende) (LAV * L, maintenant devenu bl) est une mutation qui dilue le plumage sur l’ensemble de l’oiseau et offre une large gamme de mutations de couleurs, selon la base et si c’est sous sa forme homozygote ou hétérozygote.
C’est l’une des mutations/dilutions de couleur renommées par Minvielle et al. en 2002. Le nom de cette mutation/dilution était précédemment bleu (Cheng et Kimura, 1990).
La mutation dans l’adulte ce montre tout gris bleuâtre sur le corps avec le même motif de plumage primaire que le type Sauvage.
Le motif de plumage secondaire (la marque dans chaque plume individuelle) est également le même que celui du type Sauvage.
Cependant, dans le contour individuelle des plumes, la zone de couleur brunâtre est remplacée par du gris bleuâtre et un marquage de couleur Sauvage avec un aspect blanchâtre.
Sur le type sauvage, ce gène change les marques noires en bleu-gris clair et change le rouge en chamois.
*peut également apparaître “Bleu poudre/Blanc” sous forme homozygote
Description photo: Phénotypes de type Sauvage (gauche) et Bleu (droite)
Argent
Mutation qui dilue l’ensemble du plumage à un ton argenté, mais il peut rester un peu de Fauve.
* Peut également apparaître “Ivoire/Blanc” sous forme homozygote
Lavande
Mutation qui dilue l’ensemble du plumage à un lavande/gris, mais les tons rouges restent.
Cannelle
“Cinnamon” en Anglais.
Mutation qui dilue le plumage et change la couleur des yeux en rouge.
Plumage uniforme de couleur rouge cannelle avec des yeux rouges.

OZ Snowy
Mutation découverte en Australie et pas encore disponible aux États-Unis/Europe.
Cet mutation as une teinte rouge subtile quand comparé à la mutation “Sandy”.
Crédit Photo: Tamara Rowsell d’Australie. Merci ! 🙂

OZ Sandy
Mutation découverte en Australie et pas encore disponible aux États-Unis/Europe.
Sandy a beaucoup de noir et très peu de rouge. La mutation “Snowy” est plus rouge que “Sandy”. Les nuances sont très similaires et la présence de Calico affectera également les deux de manière similaire, ce qui peut compliquer leur identification.
Crédit Photo: Tamara Rowsell d’Australie. Merci ! 🙂
“OZ-Diluer Récessif” (“OZ Dilute Recessive” en Anglais)
“OZ-Diluer Récessif” est une mutation de dilution récemment découverte qui se présente comme un oiseau gris moyen. Les mâles et les femelles présentent le motif de plumage sous-jacent et un plumage gris moyen similaire. Cette mutation semble également interagir avec de nombreuses autres mutations.
Pour les croisements:
Sauvage x Bleu = 100% Sauvage (avec un % de porteurs de gènes Bleu)
Sauvage x Bleu Hétérozygote (Brun Étendu) = 50% Sauvage, 50% Bleu Sauvage Hétérozygote
Sauvage x Bleu Homozygote (Brun Étendu) = 100% Bleu Sauvage Hétérozygote
Sauvage x Argent Sauvage = 50% Sauvage, 50% Argent Hétérozygote

Tuxedo
Comment fonctionne cette mutation basée sur le “Blanc” (“White” en Anglais) lors de la création de variétés “Tuxedo” ?
En combinant du Blanc hétérozygote avec n’importe quelle autre base et/ou motif/mutation de couleur, vous pouvez créer une variété de Tuxedo à partir de cette base! C’est aussi simple que ça !
Motif bicolore, généralement avec des ailes, un ventre, une poitrine et un visage blancs, avec des plumes foncées sur le dos et la tête. Le Tuxedo peut présenter de nombreuses couleurs de base, telles que le Tibétain, le Rosetta et le Sauvage.
Voici quelques exemples des variétés de Tuxedos les plus populaires :
Tuxedo Tibétain
Plumage brun foncé avec du Blanc
Tuxedo Rosetta
Plumage strié brun foncé et doré avec du Blanc
Tuxedo type Sauvage
Plumage de type Sauvage avec du Blanc.
Tuxedo Argenté
Plumage de type Argent avec du Blanc.
Tuxedo “German Pastel”
Plumage de type “German Pastel” avec du Blanc.
Tuxedo “Range”
Plumage brun rougeâtre avec du Blanc.
Tuxedo “Scarlet”
Plumage strié brun rougeâtre (et plus clair encore) avec du Blanc.
Pour les croisements:
Sauvage x Tuxedo = 50 % Sauvage, 50 % Tuxedo
Blanc Anglais x Tudexo = 50% Blanc Anglais, 50% Tuxedo

Autres Mutations

Albinos complet
Nom Anglais ISO: Complet Albino
Comprendre l’albinisme chez Coturnix Japonica (Caille du Japon)
L’albinisme, une maladie génétique caractérisée par un manque de pigmentation de la peau, des cheveux et des yeux, n’est pas exclusif à l’homme. Elle affecte également de nombreuses espèces animales, dont Coturnix Japonica, plus connue sous le nom de Caille du Japon.
La caille japonaise est une espèce de caille de l’Ancien Monde trouvée en Asie de l’Est, connue pour son vol rapide et son appel distinctif. Ils sont également un choix populaire dans l’aviculture et la recherche en laboratoire en raison de leur nature robuste et de leur maturité rapide. La survenue de l’albinisme chez ces oiseaux, bien que relativement rare, présente une opportunité unique d’étudier le trouble dans un contexte non humain.
Chez la Caille du Japon, l’albinisme résulte d’une mutation génétique récessive qui interfère avec la production de mélanine, le pigment responsable de la coloration distinctive de l’oiseau. Les cailles albinos présentent des plumes blanches, une peau rosâtre et des yeux rouges ou roses en raison de la réflexion de la lumière des vaisseaux sanguins derrière leurs rétines.
Ce manque de pigmentation peut rendre les cailles albinos plus vulnérables aux prédateurs et plus sensibles au soleil. Cela est dû à l’absence de mélanine, qui non seulement fournit un camouflage, mais protège également la peau et les yeux des rayons UV. Cependant, dans des conditions de captivité où ces facteurs environnementaux sont contrôlés, les cailles albinos peuvent mener une vie saine et normale.
Scientifiquement, l’albinisme chez la Caille du Japon est un domaine de recherche en cours. Comme ces oiseaux se reproduisent rapidement et ont un temps de génération court, ils sont devenus un organisme modèle précieux dans les études génétiques. Le trait d’albinisme est facile à observer et à suivre à travers les générations, ce qui fait de ces oiseaux un outil utile pour comprendre l’hérédité et l’expression de l’albinisme et d’autres conditions génétiques.
De plus, l’étude de l’albinisme chez les Cailles du Japon contribue à une compréhension plus large de l’albinisme chez les oiseaux et d’autres animaux. Les mécanismes génétiques derrière l’albinisme sont assez complexes et impliquent divers gènes, la recherche sur les cailles japonaises aidant à découvrir ces complexités.
En conclusion, l’albinisme chez la Caille du Japon est plus qu’une simple différence esthétique ; c’est une fenêtre sur le monde complexe de la génétique. L’étude de cette condition chez les cailles japonaises offre donc un grand potentiel pour améliorer notre compréhension des troubles génétiques et des principes de l’hérédité.

Diluer Mortel
Nom Anglais ISO: Dilute down lethal
Le Diluer mortel est généralement fatal dans les 1 à 3 premières semaines de vie.
(Tsudzuki, et al. 1997): Cette mutation autosomique récessive de la couleur du plumage a été trouvée dans le poussin mutant nouveau-né avec le même motif de plumage rayé que le type sauvage. Cependant, la couleur du plumage est légèrement plus claire que le type sauvage. Les rayures brunâtres du type sauvage sont remplacées par des rayures de couleur brunâtre ou jaunâtre légèrement plus claires. Dans les ailes et les cuisses du poussin mutant, des marques noires floues remplacent les marques noires denses du type sauvage. Outre l’anomalie de la couleur du duvet, les poussins mutants ont les doigts tordus.
Tous les poussins mutants meurent dans les trois jours suivant l’éclosion.

Albinisme Imparfait
Nom Anglais ISO: Imperfect albinism
Albinisme imparfait (al) et cannelle liée au sexe (al c) sont des mutations de couleur du plumage et des allèles au locus al sur le chromosome Z. L’allèle al c est dominant sur l’allèle al. Gunnarsson et coll. as constaté que le gène SLC45A2 est le gène responsable des mutations au locus al. Dans l’albinisme imparfait, une trans-version de G à T réside sur le site de l’accepteur d’épissure juste avant l’exon ing. La mutation du site d’épissage conduit à un saut d’exon de SLC45A2 et 47 acides aminés sont manquant dans la protéine mature. Gunnarson et coll. as également observé une association d’une transition de C à A à nt287 (Ala72Asp) dans l’exon 1 de SLC45A2 avec l’allèle cannelle liée au sexe (al c).
Description photo: Phénotypes de type Albinisme imparfait Sauvage (AL * N), Albinisme imparfait Cannelle (AL * C) et d’Albinisme imparfait lié au sexe (AL * A).

“Lumière Basse”
Nom Anglais ISO: Light down
La couleur de cette mutation est contrôlée par un gène autosomique dominant.
Les homozygotes, dont un tiers meurent avant l’éclosion, montre des plumules de duvet jaune crème.
L’œil des homozygotes n’est pas rougeâtre, mais le diamètre des pupilles est beaucoup plus grand que celui de l’hétérozygotes.
Par conséquent, la zone de l’iris des homozygotes est extrêmement petit.
Tous les poussins homozygotes démontrent des anomalies comportementales et d’expressivité.
Les poussins légèrement infectés secouent lentement la tête droite et gauche, tandis que les poussins gravement infectés se plient le cou vers l’intérieur, et par la suite tire vers l’arrière ou roule vers l’avant.
Certains poussins gravement infectés dresse la tête en arrière avec le bec pointé vers le haut.
Tous les poussins homozygotes qui ont éclot meurent dans la semaine suivant éclosion.
Le plumage néonatal des hétérozygotes est similaire à celle du type sauvage.
Cependant, les rayures noires sur la tête et le dos sont un peu plus étroites que celles de type sauvage, et leur couleur est légèrement plus diluée.
En conséquence, les rayures brunâtres (jaunâtres) sont plus larges que celles du type sauvage, et la couleur est également plus claire.
Le plumage entier des hétérozygotes adultes est également similaire à
celui du type sauvage, mais sa couleur est légèrement diluée sur tout le corps comme chez les poussins.
Dans le plumes individuelles des hétérozygotes adultes, le nombre de barres transversales est augmenté.
Par exemple, dans les plumes du dos, ce mutant a quatre paires de barres transversales, tandis que le type sauvage a trois paires.
Description photo: Phénotypes de type Sauvage (gauche) et Lumière basse (droite)
“Lumière Basse” Mortel
Nom Anglais ISO: Light down lethal
La couleur de cette mutation est contrôlée par un gène autosomique récessif avec une mortalité quasi totale.
Le motif du plumage néonatal de ce mutant est le même que celui du type sauvage.
Cependant, les rayures noires et feu du type sauvage sont remplacées par des rayures noires floues et bronzées diluées, respectivement.
Presque tous ces mutants qui ont réussi à éclore meurent avant trois semaines d’âge, la majorité mourant la première semaine.
De plus, la mortalité avant éclosion est élevée (37%) dans cette mutation.
Parmi 103 poussins testés avec cette mutation, seuls deux mâles ont survécu à l’âge adulte et reproduit normalement.
Le phénotype du mutant adulte est très similaire à celui du type sauvage.
Albinos complet
Nom Anglais ISO: Complet Albino
Comprendre l’albinisme chez Coturnix Japonica (Caille du Japon)
L’albinisme, une maladie génétique caractérisée par un manque de pigmentation de la peau, des cheveux et des yeux, n’est pas exclusif à l’homme. Elle affecte également de nombreuses espèces animales, dont Coturnix Japonica, plus connue sous le nom de Caille du Japon.
La caille japonaise est une espèce de caille de l’Ancien Monde trouvée en Asie de l’Est, connue pour son vol rapide et son appel distinctif. Ils sont également un choix populaire dans l’aviculture et la recherche en laboratoire en raison de leur nature robuste et de leur maturité rapide. La survenue de l’albinisme chez ces oiseaux, bien que relativement rare, présente une opportunité unique d’étudier le trouble dans un contexte non humain.
Chez la Caille du Japon, l’albinisme résulte d’une mutation génétique récessive qui interfère avec la production de mélanine, le pigment responsable de la coloration distinctive de l’oiseau. Les cailles albinos présentent des plumes blanches, une peau rosâtre et des yeux rouges ou roses en raison de la réflexion de la lumière des vaisseaux sanguins derrière leurs rétines.
Ce manque de pigmentation peut rendre les cailles albinos plus vulnérables aux prédateurs et plus sensibles au soleil. Cela est dû à l’absence de mélanine, qui non seulement fournit un camouflage, mais protège également la peau et les yeux des rayons UV. Cependant, dans des conditions de captivité où ces facteurs environnementaux sont contrôlés, les cailles albinos peuvent mener une vie saine et normale.
Scientifiquement, l’albinisme chez la Caille du Japon est un domaine de recherche en cours. Comme ces oiseaux se reproduisent rapidement et ont un temps de génération court, ils sont devenus un organisme modèle précieux dans les études génétiques. Le trait d’albinisme est facile à observer et à suivre à travers les générations, ce qui fait de ces oiseaux un outil utile pour comprendre l’hérédité et l’expression de l’albinisme et d’autres conditions génétiques.
De plus, l’étude de l’albinisme chez les Cailles du Japon contribue à une compréhension plus large de l’albinisme chez les oiseaux et d’autres animaux. Les mécanismes génétiques derrière l’albinisme sont assez complexes et impliquent divers gènes, la recherche sur les cailles japonaises aidant à découvrir ces complexités.
En conclusion, l’albinisme chez la Caille du Japon est plus qu’une simple différence esthétique ; c’est une fenêtre sur le monde complexe de la génétique. L’étude de cette condition chez les cailles japonaises offre donc un grand potentiel pour améliorer notre compréhension des troubles génétiques et des principes de l’hérédité.
Diluer Mortel
Nom Anglais ISO: Dilute down lethal
Le Diluer mortel est généralement fatal dans les 1 à 3 premières semaines de vie.
(Tsudzuki, et al. 1997): Cette mutation autosomique récessive de la couleur du plumage a été trouvée dans le poussin mutant nouveau-né avec le même motif de plumage rayé que le type sauvage. Cependant, la couleur du plumage est légèrement plus claire que le type sauvage. Les rayures brunâtres du type sauvage sont remplacées par des rayures de couleur brunâtre ou jaunâtre légèrement plus claires. Dans les ailes et les cuisses du poussin mutant, des marques noires floues remplacent les marques noires denses du type sauvage. Outre l’anomalie de la couleur du duvet, les poussins mutants ont les doigts tordus.
Tous les poussins mutants meurent dans les trois jours suivant l’éclosion.
Albinisme Imparfait
Nom Anglais ISO: Imperfect albinism
Albinisme imparfait (al) et cannelle liée au sexe (al c) sont des mutations de couleur du plumage et des allèles au locus al sur le chromosome Z. L’allèle al c est dominant sur l’allèle al. Gunnarsson et coll. as constaté que le gène SLC45A2 est le gène responsable des mutations au locus al. Dans l’albinisme imparfait, une trans-version de G à T réside sur le site de l’accepteur d’épissure juste avant l’exon ing. La mutation du site d’épissage conduit à un saut d’exon de SLC45A2 et 47 acides aminés sont manquant dans la protéine mature. Gunnarson et coll. as également observé une association d’une transition de C à A à nt287 (Ala72Asp) dans l’exon 1 de SLC45A2 avec l’allèle cannelle liée au sexe (al c).
Description photo: Phénotypes de type Albinisme imparfait Sauvage (AL * N), Albinisme imparfait Cannelle (AL * C) et d’Albinisme imparfait lié au sexe (AL * A).
“Lumière Basse”
Nom Anglais ISO: Light down
La couleur de cette mutation est contrôlée par un gène autosomique dominant.
Les homozygotes, dont un tiers meurent avant l’éclosion, montre des plumules de duvet jaune crème.
L’œil des homozygotes n’est pas rougeâtre, mais le diamètre des pupilles est beaucoup plus grand que celui de l’hétérozygotes.
Par conséquent, la zone de l’iris des homozygotes est extrêmement petit.
Tous les poussins homozygotes démontrent des anomalies comportementales et d’expressivité.
Les poussins légèrement infectés secouent lentement la tête droite et gauche, tandis que les poussins gravement infectés se plient le cou vers l’intérieur, et par la suite tire vers l’arrière ou roule vers l’avant.
Certains poussins gravement infectés dresse la tête en arrière avec le bec pointé vers le haut.
Tous les poussins homozygotes qui ont éclot meurent dans la semaine suivant éclosion.
Le plumage néonatal des hétérozygotes est similaire à celle du type sauvage.
Cependant, les rayures noires sur la tête et le dos sont un peu plus étroites que celles de type sauvage, et leur couleur est légèrement plus diluée.
En conséquence, les rayures brunâtres (jaunâtres) sont plus larges que celles du type sauvage, et la couleur est également plus claire.
Le plumage entier des hétérozygotes adultes est également similaire à
celui du type sauvage, mais sa couleur est légèrement diluée sur tout le corps comme chez les poussins.
Dans le plumes individuelles des hétérozygotes adultes, le nombre de barres transversales est augmenté.
Par exemple, dans les plumes du dos, ce mutant a quatre paires de barres transversales, tandis que le type sauvage a trois paires.
Description photo: Phénotypes de type Sauvage (gauche) et Lumière basse (droite)
“Lumière Basse” Mortel
Nom Anglais ISO: Light down lethal
La couleur de cette mutation est contrôlée par un gène autosomique récessif avec une mortalité quasi totale.
Le motif du plumage néonatal de ce mutant est le même que celui du type sauvage.
Cependant, les rayures noires et feu du type sauvage sont remplacées par des rayures noires floues et bronzées diluées, respectivement.
Presque tous ces mutants qui ont réussi à éclore meurent avant trois semaines d’âge, la majorité mourant la première semaine.
De plus, la mortalité avant éclosion est élevée (37%) dans cette mutation.
Parmi 103 poussins testés avec cette mutation, seuls deux mâles ont survécu à l’âge adulte et reproduit normalement.
Le phénotype du mutant adulte est très similaire à celui du type sauvage.
Albinos complet
Nom Anglais ISO: Complet Albino
Comprendre l’albinisme chez Coturnix Japonica (Caille du Japon)
L’albinisme, une maladie génétique caractérisée par un manque de pigmentation de la peau, des cheveux et des yeux, n’est pas exclusif à l’homme. Elle affecte également de nombreuses espèces animales, dont Coturnix Japonica, plus connue sous le nom de Caille du Japon.
La caille japonaise est une espèce de caille de l’Ancien Monde trouvée en Asie de l’Est, connue pour son vol rapide et son appel distinctif. Ils sont également un choix populaire dans l’aviculture et la recherche en laboratoire en raison de leur nature robuste et de leur maturité rapide. La survenue de l’albinisme chez ces oiseaux, bien que relativement rare, présente une opportunité unique d’étudier le trouble dans un contexte non humain.
Chez la Caille du Japon, l’albinisme résulte d’une mutation génétique récessive qui interfère avec la production de mélanine, le pigment responsable de la coloration distinctive de l’oiseau. Les cailles albinos présentent des plumes blanches, une peau rosâtre et des yeux rouges ou roses en raison de la réflexion de la lumière des vaisseaux sanguins derrière leurs rétines.
Ce manque de pigmentation peut rendre les cailles albinos plus vulnérables aux prédateurs et plus sensibles au soleil. Cela est dû à l’absence de mélanine, qui non seulement fournit un camouflage, mais protège également la peau et les yeux des rayons UV. Cependant, dans des conditions de captivité où ces facteurs environnementaux sont contrôlés, les cailles albinos peuvent mener une vie saine et normale.
Scientifiquement, l’albinisme chez la Caille du Japon est un domaine de recherche en cours. Comme ces oiseaux se reproduisent rapidement et ont un temps de génération court, ils sont devenus un organisme modèle précieux dans les études génétiques. Le trait d’albinisme est facile à observer et à suivre à travers les générations, ce qui fait de ces oiseaux un outil utile pour comprendre l’hérédité et l’expression de l’albinisme et d’autres conditions génétiques.
De plus, l’étude de l’albinisme chez les Cailles du Japon contribue à une compréhension plus large de l’albinisme chez les oiseaux et d’autres animaux. Les mécanismes génétiques derrière l’albinisme sont assez complexes et impliquent divers gènes, la recherche sur les cailles japonaises aidant à découvrir ces complexités.
En conclusion, l’albinisme chez la Caille du Japon est plus qu’une simple différence esthétique ; c’est une fenêtre sur le monde complexe de la génétique. L’étude de cette condition chez les cailles japonaises offre donc un grand potentiel pour améliorer notre compréhension des troubles génétiques et des principes de l’hérédité.
Diluer Mortel
Nom Anglais ISO: Dilute down lethal
Le Diluer mortel est généralement fatal dans les 1 à 3 premières semaines de vie.
(Tsudzuki, et al. 1997): Cette mutation autosomique récessive de la couleur du plumage a été trouvée dans le poussin mutant nouveau-né avec le même motif de plumage rayé que le type sauvage. Cependant, la couleur du plumage est légèrement plus claire que le type sauvage. Les rayures brunâtres du type sauvage sont remplacées par des rayures de couleur brunâtre ou jaunâtre légèrement plus claires. Dans les ailes et les cuisses du poussin mutant, des marques noires floues remplacent les marques noires denses du type sauvage. Outre l’anomalie de la couleur du duvet, les poussins mutants ont les doigts tordus.
Tous les poussins mutants meurent dans les trois jours suivant l’éclosion.
Albinisme Imparfait
Nom Anglais ISO: Imperfect albinism
Albinisme imparfait (al) et cannelle liée au sexe (al c) sont des mutations de couleur du plumage et des allèles au locus al sur le chromosome Z. L’allèle al c est dominant sur l’allèle al. Gunnarsson et coll. as constaté que le gène SLC45A2 est le gène responsable des mutations au locus al. Dans l’albinisme imparfait, une trans-version de G à T réside sur le site de l’accepteur d’épissure juste avant l’exon ing. La mutation du site d’épissage conduit à un saut d’exon de SLC45A2 et 47 acides aminés sont manquant dans la protéine mature. Gunnarson et coll. as également observé une association d’une transition de C à A à nt287 (Ala72Asp) dans l’exon 1 de SLC45A2 avec l’allèle cannelle liée au sexe (al c).
Description photo: Phénotypes de type Albinisme imparfait Sauvage (AL * N), Albinisme imparfait Cannelle (AL * C) et d’Albinisme imparfait lié au sexe (AL * A).
“Lumière Basse”
Nom Anglais ISO: Light down
La couleur de cette mutation est contrôlée par un gène autosomique dominant.
Les homozygotes, dont un tiers meurent avant l’éclosion, montre des plumules de duvet jaune crème.
L’œil des homozygotes n’est pas rougeâtre, mais le diamètre des pupilles est beaucoup plus grand que celui de l’hétérozygotes.
Par conséquent, la zone de l’iris des homozygotes est extrêmement petit.
Tous les poussins homozygotes démontrent des anomalies comportementales et d’expressivité.
Les poussins légèrement infectés secouent lentement la tête droite et gauche, tandis que les poussins gravement infectés se plient le cou vers l’intérieur, et par la suite tire vers l’arrière ou roule vers l’avant.
Certains poussins gravement infectés dresse la tête en arrière avec le bec pointé vers le haut.
Tous les poussins homozygotes qui ont éclot meurent dans la semaine suivant éclosion.
Le plumage néonatal des hétérozygotes est similaire à celle du type sauvage.
Cependant, les rayures noires sur la tête et le dos sont un peu plus étroites que celles de type sauvage, et leur couleur est légèrement plus diluée.
En conséquence, les rayures brunâtres (jaunâtres) sont plus larges que celles du type sauvage, et la couleur est également plus claire.
Le plumage entier des hétérozygotes adultes est également similaire à
celui du type sauvage, mais sa couleur est légèrement diluée sur tout le corps comme chez les poussins.
Dans le plumes individuelles des hétérozygotes adultes, le nombre de barres transversales est augmenté.
Par exemple, dans les plumes du dos, ce mutant a quatre paires de barres transversales, tandis que le type sauvage a trois paires.
Description photo: Phénotypes de type Sauvage (gauche) et Lumière basse (droite)
“Lumière Basse” Mortel
Nom Anglais ISO: Light down lethal
La couleur de cette mutation est contrôlée par un gène autosomique récessif avec une mortalité quasi totale.
Le motif du plumage néonatal de ce mutant est le même que celui du type sauvage.
Cependant, les rayures noires et feu du type sauvage sont remplacées par des rayures noires floues et bronzées diluées, respectivement.
Presque tous ces mutants qui ont réussi à éclore meurent avant trois semaines d’âge, la majorité mourant la première semaine.
De plus, la mortalité avant éclosion est élevée (37%) dans cette mutation.
Parmi 103 poussins testés avec cette mutation, seuls deux mâles ont survécu à l’âge adulte et reproduit normalement.
Le phénotype du mutant adulte est très similaire à celui du type sauvage.

Article rédigé et utilisant des sources traduites de l’Anglais par
Boris Nicoloff pour Bisou ma Caille et Master Caille
Droits de reproduction et de diffusion réservés ©Bisou ma Caille / Master Caille
Sources et crédits des articles utilisés dans cet article:
Gunnarsson U, Hellström AR, Tixier-Boichard M, et al. Mutations in SLC45A2 cause plumage color variation in chicken and Japanese quail. Genetics. 2007;175(2):867-877. doi:10.1534/genetics.106.063107
Minvielle F, Gourichon D and Monvoisin JL. Testing homology of loci for two plumage colors, “lavender” and “recessive white”, with chicken and Japanese quail hybrids. The Journal of Heredity, 93: 73-76, 2002
Bed’hom, B., Vaez, M., Coville, J. et al. The lavender plumage colour in Japanese quail is associated with a complex mutation in the region of MLPH that is related to differences in growth, feed consumption and body temperature. BMC Genomics 13, 442 (2012). https://doi.org/10.1186/1471-2164-13-442
Tsudzuki, Masaoki. (2008). Mutations of Japanese Quail (Coturnix japonica) and Recent Advances of Molecular Genetics for This Species. Journal of Poultry Science – J POULT SCI. 45. 159-179. 10.2141/jpsa.45.159.
Sources et crédits des photos utilisés dans cet article:
“OZ Sandy” et “OZ Snowy”: Tamara Rowsell d’Australie. Merci ! 🙂
“Silver”: Cora Guindon, USA. Merci ! 🙂

